










Introduction
In general, peptides have a smaller molecular structure than proteins, consisting of 2–50 amino acids. Certain peptides play a role in regulating the activity of other molecules. Bioactive peptides consist of 2–20 amino acids and have a relatively small molecular weight compared to proteins (Lafarga and Hayes, 2014). The market for bioactive peptides is expanding with the growth of functional food and beverage products, and they are widely applied in functional foods, natural health products, health foods, and cosmetics (Chalamaiah et al., 2019). This growth can be attributed to the fact that consumers are becoming more health-conscious, and industries are utilizing functional ingredients to develop new products. Bioactive peptides are found in food proteins, especially in milk, meat, fish, and legumes. They are utilized as ingredients in functional foods and pharmaceuticals due to their beneficial effects on the human digestive, endocrine, nervous, and cardiovascular systems, among others, and their role in health (Heres et al., 2021a; Sánchez and Vázquez, 2017). The efficacy of bioactive peptides is often determined by their molecular weight and amino acid sequence because the amino acids that comprise the peptide sequence can have varying properties and effects. Livestock-derived bioactive peptides have been reported to have antioxidant, antihypertensive, antithrombotic, and antimicrobial activities, which have positive effects on disease prevention and blood circulation (Aluko, 2015; Kim et al., 2024; Lafarga and Hayes, 2014; Rubak et al., 2022). Previous studies have confirmed that bioactive peptides that modulate various biological actions can be obtained from pork (Arihara, 2006). In particular, antioxidant and antimicrobial active peptides isolated from pork muscle proteins provide important health benefits to humans and can be utilized as functional ingredients in foods (Di Bernardini et al., 2011). However, the effective utilization of pork-derived bioactive peptides has not been adequately studied. Therefore, in this study, we introduced a method for protein digestion analysis that can be utilized to obtain peptides and categorize various potencies and types of bioactive peptides derived from pork.
Process of Protein Digestion by Enzymes
Proteins consumed by humans must be hydrolyzed by proteolytic enzymes secreted by the stomach, pancreas, and small intestine in order to be digested and absorbed by the body. After proteins digestion, peptides present in the intestinal lumen typically consist of 2–6 amino acids, which account for about 80% of the total amino acids (Bhutia and Ganapathy, 2018). In the intestinal lumen, the amount of amino acids present in peptides is higher than that of free amino acids (Adibi and Mercer, 1973). Consequently, most peptides and free amino acids are transported across the intestinal epithelium into the digestive tract through the brush border membrane transport system. The majority of peptides are then hydrolyzed to free amino acids, which make up about 90% of the total amino acids (Bhutia and Ganapathy, 2018).
Ingested proteins are broken down by a variety of enzymes secreted by the body’s digestive system. The initial step in this process is performed by pepsin, a proteolytic enzyme secreted by the stomach. Pepsin is initially secreted as an inactive precursor, called pepsinogen, which is produced by the chief cells of the stomach (Gupta, 2018). This inactive precursor, pepsinogen, is then activated through an autocatalytic reaction in the acidic pH environment of the stomach, resulting in the production of pepsin (Gupta, 2018). Protein hydrolysates processed by pepsin are mostly in the form of polypeptides, and only a small amount of free amino acids is released through hydrolysis (Hinsberger and Sandhu, 2004). After undergoing digestion in the stomach, gastric contents pass through the duodenum and jejunum, where they stimulate cells in the intestinal mucosa to produce cholecystokinin (Liddle, 1997). Cholecystokinin then triggers the secretion of pancreatic juice, which is rich in proteolytic enzymes, and causes the gallbladder to contract and release bile (Liddle, 1997). In addition, when the gastric contents reach the small intestine, the acidic pH environment created by gastric acid prompts S-cells in the duodenum to release secretin, which is produced by these cells (DiGregorio and Sharma, 2019). Secretin increases the secretion of bicarbonate from the pancreas and biliary tract. This neutralizes the acidic pH environment in the duodenum caused by stomach acid to a pH level of around 6–8 and reduces the secretion of stomach acid (Bhutia and Ganapathy, 2018; DiGregorio and Sharma, 2019).
The pancreas is a vital digestive organ that produces and secretes proteolytic enzymes into the small intestine to digest ingested protein. The major pancreatic proteolytic enzymes have been identified as trypsin, chymotrypsin, elastase, and carboxypeptidase (Whitcomb and Lowe, 2007). Similar to pepsin, these enzymes are initially secreted as inactive precursors, including trypsinogen, chymotrypsinogen, proelastase, and procarboxypeptidase (Whitcomb and Lowe, 2007). Among them, trypsinogen is first activated to trypsin by enteropeptidase in the small intestine. Activated trypsin then acts on chymotrypsinogen, proelastase, and procarboxypeptidase to form active chymotrypsin, elastase, and carboxypeptidase (Bhutia and Ganapathy, 2018). Trypsin is highly reactive towards peptides containing the basic amino acids arginine and lysine, while chymotrypsin hydrolyzes peptides containing the aromatic amino acids tyrosine, phenylalanine, and tryptophan (Whitcomb and Lowe, 2007). In addition, elastase acts on peptide binding sites formed by the non-polar amino acids glycine and alanine (Whitcomb and Lowe, 2007). Consequently, the proteolytic enzymes in the pancreas hydrolyze proteins that are mostly present in polypeptide form during small intestinal digestion into oligopeptides and free amino acids consisting of 6–8 amino acids (Bhutia and Ganapathy, 2018). These oligopeptides are then hydrolyzed into smaller forms of peptides, such as tripeptides and dipeptides, by brush border peptidases found in the microvilli composed of small intestinal enterocytes (Hooton et al., 2015). Finally, the myriad of tripeptides and dipeptides are absorbed into the small intestine where they are broken down into amino acids by cytoplasmic peptidases and released into the bloodstream (Boron and Boulpaep, 2016).
Analysis Methods for Protein Digestibility
Methods such as the bicinchoninic acid (BCA) assay, sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), gel permeation chromatography (GPC), and in vitro digestion system have been utilized to analyze protein digestibility and molecular weight distribution, which are relevant for peptide acquisition (Li et al., 2017; Rezvankhah et al., 2021; Wen et al., 2015).
The BCA assay is a highly sensitive method for quantifying proteins by comparing their chromogenic reactions. The principle behind this method is that Cu2+ ions are reduced to Cu+ ions by peptide binding of proteins in an alkaline environment. The Cu+ ions then combine with BCA to form a purple complex. This assay is similar to the Lowry assay but has the advantage of being relatively simple and resistant to compounds that may interfere with the results (Walker, 2009). Once the reaction is complete, the complex can be analyzed using a spectrophotometer to measure the amount of protein at the maximum absorption wavelength of 562 nm (He, 2011). This reaction is mainly influenced by the presence of four amino acid residues (cysteine, cystine, tyrosine, and tryptophan) in the protein molecule (Fischer et al., 1999). Therefore, the BCA assay can be utilized to determine the initial protein content of a sample.
SDS-PAGE is an electrophoresis technique that analyzes the movement of charged protein molecules in an electric field. It is commonly used to separate proteins by size and analyze them qualitatively (Rajput and Sharma, 2011; Roy et al., 2012). SDS, an anionic surfactant with a strong protein denaturing effect, binds to proteins at a constant rate. During this process, the proteins are transformed into a linear chain structure and become negatively charged (Farrell, 2009; Rajput and Sharma, 2011). The proteins then move in the electric field containing the polyacrylamide gel according to their molecular size, with smaller proteins migrating and separating faster than larger proteins (Rajput and Sharma, 2011). Therefore, SDS-PAGE separates polypeptides based on molecular size, making it the best experimental method for analyzing protein digestibility as a function of protein molecular weight (Righetti, 2005). Li et al. (2017) and Wen et al. (2015) used SDS-PAGE to screen for changes in molecular weight and compare the digestibility of pork protein before and after in vitro digestion. Specifically, Li et al. (2017) compared the molecular weight of four types of pork proteins (cooked, emulsion-type sausage, dry-cured, and stewed) before digestion, after pepsin digestion, and after pepsin and trypsin digestion. Additionally, Wen et al. (2015) compared the differences in molecular weight of proteins from four types of cooked meat (pork, beef, chicken, and fish) before digestion, after pepsin digestion, and after pepsin and trypsin digestion. In both studies, compared to undigested samples, samples treated with pepsin alone lost protein bands greater than 150 kDa, and a relatively greater amount of protein bands between 50–100 kDa was identified (Li et al., 2017; Wen et al., 2015). These results indicate that high molecular weight proteins were digested and decomposed into small sizes. In samples treated with pepsin and trypsin together, proteins were decomposed more effectively, and the degraded proteins were confirmed to be clustered in the range between 2–10 kDa (Li et al., 2017; Wen et al., 2015).
Size exclusion chromatography (SEC), a chromatography method developed in 1955, is the most commonly method used to separate polymers such as proteins and peptides according to their molecular size. SEC is utilized for various purposes including adsorption, desalting, and determining molecular weight distribution (Sorci and Belfort, 2014; Wang et al., 2017). As the polymer moves through the column, larger molecules elute faster because they cannot penetrate the pores of the gel, while smaller molecules can penetrate the pores and move freely, increasing the elution time (Boone and Adamec, 2016; Deb et al., 2019). SEC is often used interchangeably with GPC or gel filtration chromatography. GPC is a method of separating molecules by size through elution from a column composed of porous gels such as dextran, agarose, and polyacrylamide. This method can be utilized for extracting specific proteins and analyzing the molecular weight distribution of hydrolysates (Jia et al., 2010; Lee et al., 2022; Ting et al., 2013).
Protein digestibility can be assessed through both in vivo and in vitro experiments. Kjeldahl assay can be used after feeding experimental animals to determine the crude protein content in feed samples and feces, as well as to calculate feed intake for examining the digestibility of apparent proteins and peptides (AOAC, 2000; Kumar et al., 2019). In vivo digestion experiments can provide the most accurate results, but they are time-consuming, costly, and subject to ethical constraints (Boisen and Eggum, 1991; Guerra et al., 2012).
In vitro digestion simulation systems are more efficient compared to in vivo digestion experiments and are widely used to evaluate protein digestibility and physiological properties. In vitro models have been proposed as an alternative to the financial and ethical challenges of in vivo experiments involving humans or animals (Bohn et al., 2018). These systems can be utilized to rapidly screen various food structures and ingredients. In particular, meat (18%) has been identified as the most commonly studied food product using in vitro digestion simulation systems after plant foods (45%; Coles et al., 2005; Hur et al., 2011). In a previous study, an in vitro digestion simulation system was utilized to identify biochemical indicators of digestibility and nutritional properties of pork muscle protein following different meat processing methods (Bax et al., 2012). Lee et al. (2020) investigated the digestibility and antioxidant properties of beef protein according to aging period and cooking conditions by simulating the digestive conditions of infants. Gallego et al. (2020) also employed an in vitro digestion simulation system to evaluate the antioxidant activity of peptides detected after digestion in dry-brined pork hindquarters. However, further testing is needed to confirm the similarity of results obtained from studies using these in vitro digestion simulation systems when applied to in vivo models.
Utilizing an in vitro digestion mimicry system, researchers have identified peptides in fibrillar protein hydrolysates from porcine loin muscle that exhibit partial sequence homology to peptides found through in vivo digestion experiments (Escudero et al., 2010b). For example, the peptide AGDDAPR, identified in pork actin, has been found to share partial sequence homology with AGDDAPRAVF and AGFAGDDAPR identified in the duodenum or jejunum of pigs after consuming beef and trout (Bauchart et al., 2007; Escudero et al., 2010b). However, the digestive enzymes, conditions, and other factors in for each stage of digestion in the in vitro simulation system can vary based on age and sex, making it challenging to replicate results from in vivo animal experiments. Therefore, comprehensive research is needed to achieve similar outcomes as in vivo experiments.
The Applicable Bioavailability Methods for Bioactive Peptides
The antioxidant activity of proteins and peptides is manifested through mechanisms such as free radical scavenging, inactivation of reactive oxygen species, chelation of metal ion, and antioxidant enzyme activity (Elias et al., 2008; Yan et al., 2020).
Free radicals are atoms, molecules, or ions that possess an unpaired electron, making them unstable and highly reactive with other organic compounds (Lobo et al., 2010). These free radicals and other reactive oxygen species derived from oxygen are generated in the body through normal cell metabolism or exposure to external factors such as smoking, ultraviolet light, ozone, and X-rays (Bagchi and Puri, 1998; Carocho and Ferreira, 2013). Reactive oxygen species include the free radicals superoxide ion (O2–), hydroxyl radical (HO), hydroperoxyl radical (HO2), and nitric oxide (NO), as well as other reactive oxygen species such as singlet oxygen (O2), hydrogen peroxide (H2O2), peroxynitrite (ONOO–), and hypochlorous acid (HClO; Carocho and Ferreira, 2013; Lobo et al., 2010). These reactive species are neutralized by antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase, and various antioxidants (Rock et al., 1996). However, when the balance between reactive species and antioxidants is disrupted, the overabundance of reactive species causes oxidative stress (Rock et al., 1996). Free radicals and other reactive oxygen species exhibit high reactivity with most cellular molecules, including amino acids, sugars, and lipids, causing cellular damage and disruption of homeostasis (Lobo et al., 2010; Young and Woodside, 2001). In addition, excessive oxidative stress can contribute to the development of cancer, liver, kidney, cardiovascular, and neurodegenerative diseases (Carocho and Ferreira, 2013; Soomro, 2019; Tönnies and Trushina, 2017). Chelation is the formation of chelate compounds through the coordination bonding of organic substances with metal ions such as iron and copper (van Lith and Ameer, 2016). Metal ions can trigger redox reactions, leading to oxidative stress and the generation of free radicals that damage biomolecules (van Lith and Ameer, 2016; Yan et al., 2020). Furthermore, an imbalance of metal ions such as iron, copper, zinc, and calcium, along with oxidative stress, can contribute to the development of Alzheimer’s disease, a neurodegenerative condition (Wang et al., 2020). Therefore, it is important for antioxidants to effectively inhibit and reduce the interactions of reactive oxygen species with DNA, proteins, lipids, and sugars.
Proteins represent the three-dimensional structure of polypeptides, and most peptides with antioxidant activity are located inside this structure. Therefore, disrupting the three-dimensional structure of proteins through methods such as heat treatment can increase the solvent accessibility of amino acid residues in peptides, thereby enhancing their antioxidant activities (Elias et al., 2008). Furthermore, enzymatic hydrolysis can increase antioxidant activity by breaking peptide bonds to expose amino acid residues. Studies have shown that the antioxidant activity of enzymatically hydrolyzed peptides is higher than that of undigested proteins (Elias et al., 2008; Park and Chin, 2011). In addition, proteins lacking metal ion storage or transport capabilities can chelate metal ions. Proteins with exposed histidine, glutamic acid, and aspartic acid on their surface have been shown to chelate metal ions (Elias et al., 2008). Therefore, there is a need for chelators derived from natural sources with minimal side effects that can bind to these metal ions to form chelate compounds. Peptides derived from pork skeletal muscle proteins have been found to chelate ferrous ions (Fe2+), and their chelating ability is enhanced through in vitro digestion (Zhu et al., 2016). Previous studies have also confirmed that peptides derived from pork proteins and collagen can chelate metal ions (Li et al., 2007; Saiga et al., 2003; Xing et al., 2016).
Experimental methods for measuring the antioxidant activity of proteins and peptides include the 2,2-azinobis-3-ethyl-benzothiazoline-6-sulfonic acid (ABTS) radical scavenging assay, 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging assay, iron-chelating assay, and reducing power assay (Acharya, 2017; Bhalodia et al., 2013; Zhong and Shahidi, 2015).
The ABTS assay is considered more responsive and less sensitive to pH than the DPPH assay. Additionally, ABTS assay saves time, money, and sample volume, making it a more efficient option (Moniruzzaman et al., 2012; Shalaby and Shanab, 2013).
The DPPH assay is widely used to evaluate antioxidant activity and is an electron transfer-based assay (Huang et al., 2005; Zhong and Shahidi, 2015). DPPH is a stable nitrogen radical with a dark purple color. Unlike the ABTS assay, the DPPH assay does not require the generation of radicals before conducting the test (Prior et al., 2005).
Iron-chelating assays are used for analyzing antioxidant activity by measuring the capacity of proteins and peptides to chelate Fe2+. This is determined by the level of chromogenicity, as proteins and peptides chelate Fe2+ to form chelate compounds, and ferrozine binds to the unchelated Fe2+ (Santos et al., 2017). Therefore, low chromogenicity indicates a strong ability of proteins and peptides to bind and chelate Fe2+, indicating high antioxidant activity (Gülçin, 2005).
The reducing power assay is another method that can be utilized to measure the antioxidant activity of proteins and peptides. Since antioxidants also function as reducing agents, reducing power is an important indicator of antioxidant activity (Shahidi and Zhong, 2015). This assay measures reducing power by detecting the reaction of a substance with potassium ferricyanide to form potassium ferrocyanide, which then reacts with ferric chloride to form a ferric-ferrous complex. This reaction results in a yellowish discoloration as the ferric form of potassium ferricyanide is reduced to the ferrous form (Bhalodia et al., 2013; Park et al., 2015).
Experimental methods for measuring antioxidant enzyme activity in proteins and peptides in vitro include the SOD assay, CAT assay, and peroxidase (POD) assay (Dasgupta and Klein, 2014; Haida and Hakiman, 2019).
SOD, an antioxidant enzyme, plays an important role in protecting biomolecules from oxidative stress induced by reactive oxygen species (Boguszewska et al., 2010). The SOD assay measures the activity of SOD, which catalyzes the conversion of the free radical superoxide ion (O2) into hydrogen peroxide (H2O2) and singlet oxygen (O2; McCord and Fridovich, 1969).
CAT, another antioxidant enzyme, is found in most tissues including the liver and stomach of animals. The CAT assay measures the activity of CAT, which catalyzes the conversion of H2O2 to O2 and H2O (Liu and Kokare, 2017; Miranda-Bautista et al., 2017). CAT can inhibit cellular damage caused by oxidative stress by reducing the amount of H2O2, a reactive oxygen species produced in vivo (Catalán et al., 2018). This is based on the principle that the amount of resorufin, a product that is reduced to O2 and H2O by CAT, and the remaining H2O2 reacts with horseradish peroxidase (HRP) and a non-fluorescent probe, which is then analyzed by fluorescence or absorbance measurements (Pinto et al., 2011). Similarly, the POD assay determines the activity of POD by measuring the amount of resorufin produced by the reaction of H2O2 with HRP and a non-fluorescent probe. Therefore, these antioxidant assays can be used to predict the mechanism of antioxidant activity of bioactive peptides by considering their principles.
The antihypertensive activity of proteins and peptides can be measured through the angiotensin converting enzyme (ACE) inhibitory assay. ACE plays a crucial role in the renin-angiotensin system (RAS), which regulates blood pressure (Gurley and Coffman, 2007; Fig. 1). Gurley and Coffman (2007) have shown that renin in the blood converts angiotensinogen produced by the liver to angiotensin I, which is then converted to angiotensin II by ACE. Within the RAS, ACE converts angiotensin I, an inactive decapeptide, to angiotensin II, an octapeptide with vasoconstrictor activity, and inactivates bradykinin, which exhibits vasodilator activity (Mora et al., 2018; Zhuo et al., 2013). Angiotensin II binds to two G protein coupled-receptors, the AT1 receptor and AT2 receptor, to carry out its biological functions (Wu et al., 2017). The AT1 receptor is associated with a variety of physiological functions, including vasoconstriction, secretion of hormones such as noradrenalin and aldosterone, and sodium reabsorption, while the AT2 receptor promotes vasodilation and sodium excretion (Carey, 2017; Contreras et al., 2003; Kaschina and Unger, 2003; Wu et al., 2017). In this context, antihypertensive functional peptides can reduce angiotensin II production by inhibiting ACE and lower blood pressure by blocking the AT1 receptor (Contreras et al., 2003; Ferrario et al., 2005). In addition, antihypertensive functional peptides play a role in balancing the vasoconstrictor and dilator effects of angiotensin I and bradykinin (Mora et al., 2018).
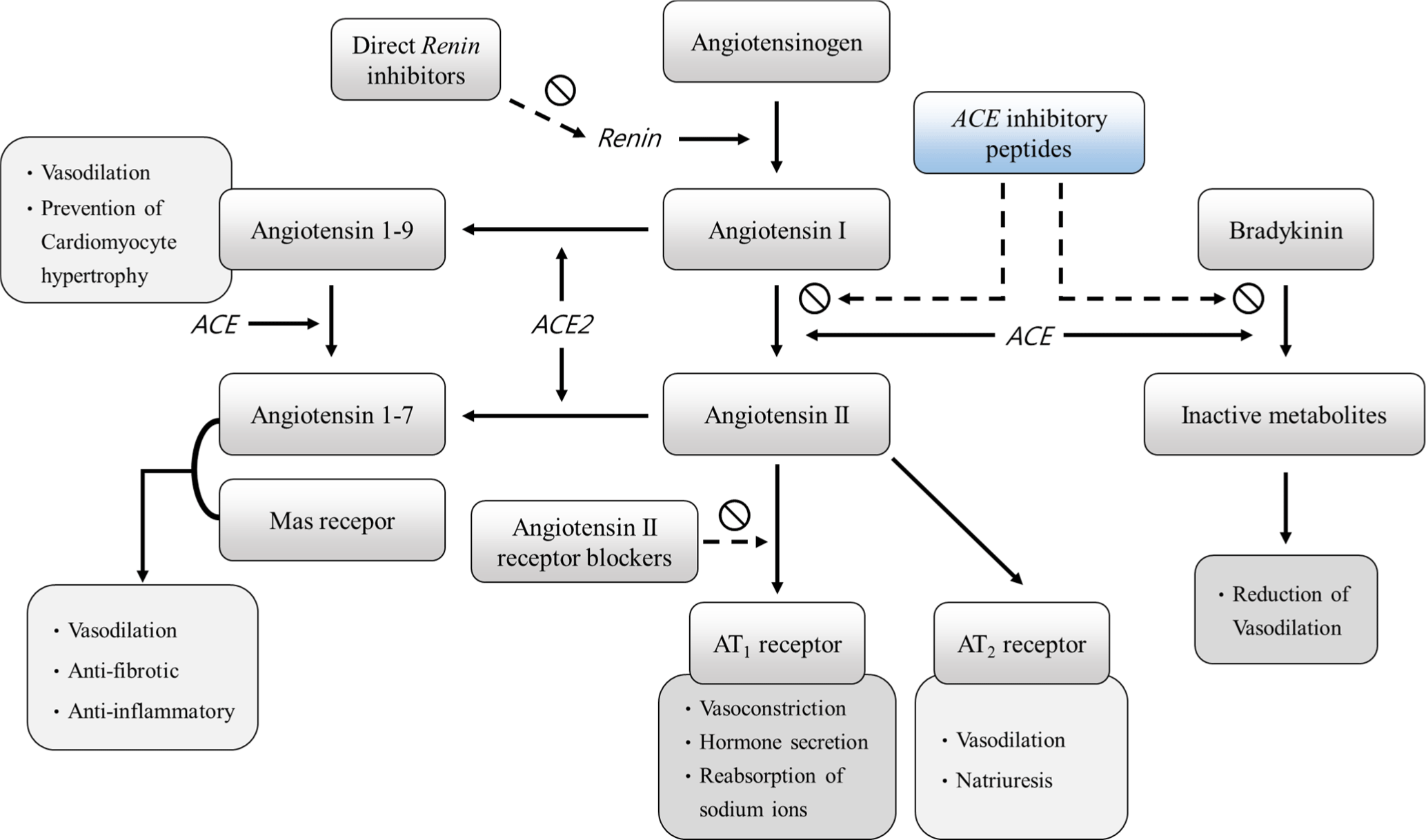
On the other hand, ACE2, which has been identified as a homologue of ACE, is known to play a physiological role in the regulation of homeostasis (Turner, 2015). In addition, ACE2 cleaves the amino acids at the C-terminus of angiotensin II to form angiotensins 1–7, which bind to the Mas receptor and exert anti-inflammatory, vasodilatory, and antifibrotic effects (Barroso et al., 2015; Shenoy et al., 2015; Simões e Silva et al., 2013). Similarly, ACE2 can hydrolyze angiotensin I to produce angiotensin 1–9, which can be converted to angiotensin 1–7 by ACE action (Donoghue et al., 2000). Previous studies have reported that angiotensin 1–9 can exhibit vasodilatory functions, reducing blood pressure and preventing cardiomyocyte hypertrophy (Gonzalez et al., 2018; Sotomayor-Flores et al., 2020). Therefore, the antihypertensive activity of the peptide may be mainly determined by ACE inhibition.
Bioactive Peptides in Pork
It has been confirmed that bioactive peptides exhibit little bioactivity in their normal protein-bound state, and their activity is triggered by protein degradation through processes such as ripening, fermentation, enzymatic hydrolysis, and digestion (Arihara and Ohata, 2008; Xing et al., 2019). Previous studies have shown that plant-derived bioactive peptides are extracted using digestive enzymes such as trypsin, chymotrypsin, and pepsin, or plant-derived proteolytic enzymes papain, bromelain, and ficin (Ryan et al., 2011; Singh et al., 2019). Additionally, alkaline proteases from microbial fermentation processes have been identified to be used to produce highly nutritious protein hydrolysates (Sharma et al., 2019; Sumantha et al., 2006). The use of commercialized proteolytic enzymes, including Protamex® and Flavourzyme®, for the production of ACE inhibitory active peptides has been previously documented (Mirdhayati et al., 2016). Furthermore, it has been reviewed that bioactivities, such as antioxidant and ACE inhibitory activities of protein hydrolysates formed by using alcalase with other proteolytic enzymes are increased (Tacias-Pascacio et al., 2020). In addition, large amounts of bioactive peptides have been produced from pork after in vitro digestion, confirming that pork can be a major source of bioactive peptides (Escudero et al., 2010b). The process of bioactive peptide formation from proteins in meat is shown in Fig. 2.
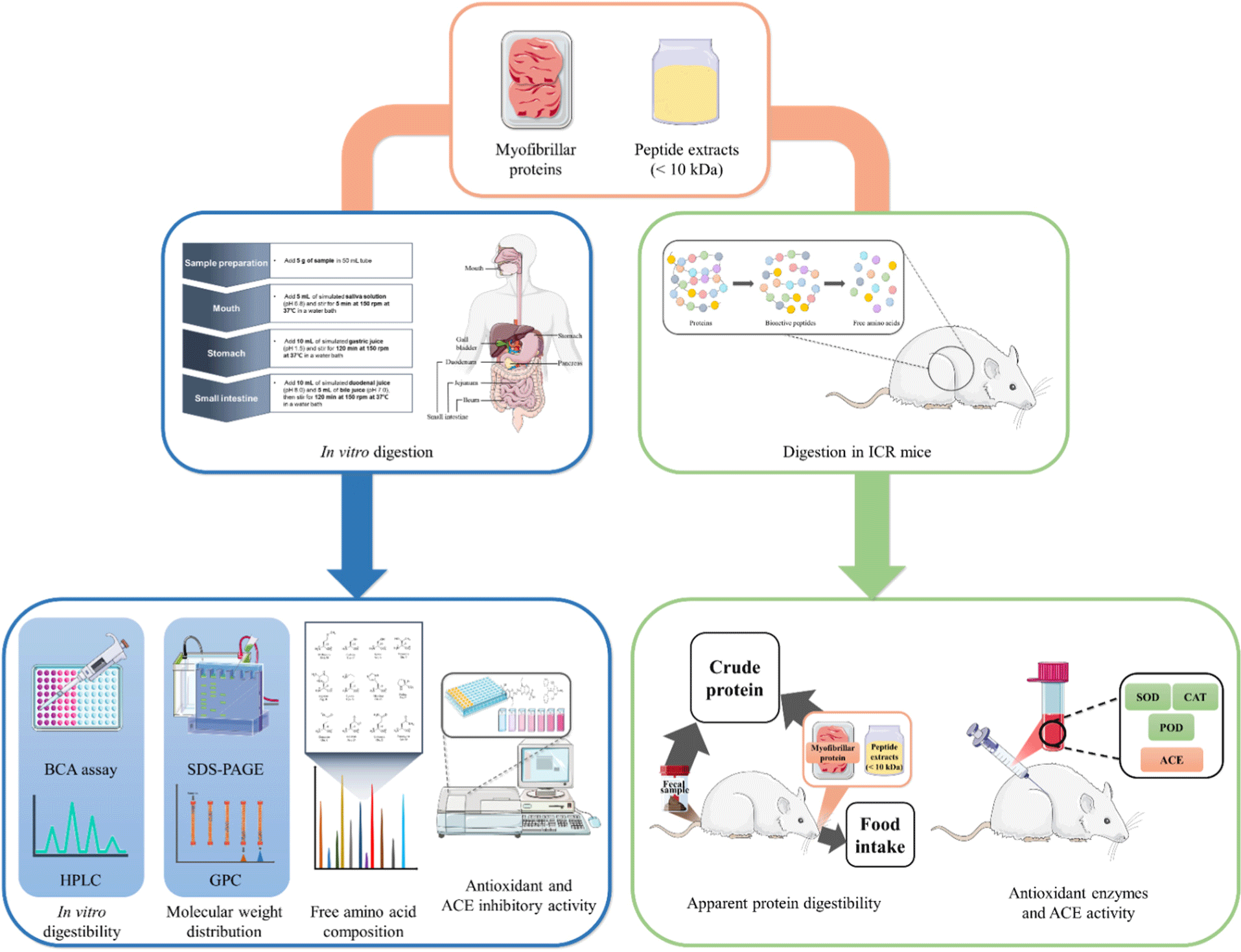
Table 1 displays the peptides with antioxidant activity derived from porcine proteins. All of these peptides have a small molecular weight, mostly less than 1 kDa. In addition, bioactive peptides with high DPPH radical scavenging and metal ion chelating activities were extracted from protein hydrolysate obtained from pork source fiber protein (Saiga et al., 2003). Carnosine and anserine, representative peptides with antioxidant activity, were also obtained from porcine loin muscle (Simonetti et al., 2019). Furthermore, bioactive peptides predicted to be generated after hydrolysis from pork myofibrillar proteins were identified through in silico analysis as potentially exhibiting a variety of bioactivities, including antioxidant, antihypertensive, antithrombotic, and dipeptidyl peptidase-IV (DPP-IV) inhibition (Keska and Stadnik, 2017). DPP-IV is an enzyme that degrades incretin, a blood sugar-regulating hormone released when food is consumed. Inhibition of DPP-IV increases incretin content, stimulating the release of insulin and inhibiting the release of glucagon, which regulates blood sugar (Drucker, 2007). Peptides extracted from dry-cured pork ham with a molecular weight of less than 1 kDa exhibit the highest antioxidant activity (Xing et al., 2018). Meanwhile, meat-derived bioactive peptides are considered to have higher antioxidant activity as they contain more hydrophobic amino acids (leucine, isoleucine, and valine) and aromatic amino acids (tryptophan, tyrosine, and phenylalanine; Peighambardoust et al., 2021).
| Protein source | Peptide sequence | Treatment | MW (Da)1) | Reference |
|---|---|---|---|---|
| Porcine myofibrillar protein (actin) | DSGVT | Papain | 650.3 | Saiga et al. (2003) |
| Porcine myofibrillar protein (unknown) | IEAEGE | Papain | 646.4 | Saiga et al. (2003) |
| Porcine myofibrillar protein (tropomyosin) | DAQEKLE | Papain | 832.5 | Saiga et al. (2003) |
| Porcine myofibrillar protein (tropomyosin) | EELDNALN | Papain | 916.9 | Saiga et al. (2003) |
| Porcine myofibrillar protein (myosin heavy chain) | VPSIDDQEELM | Papain | 1,275.0 | Saiga et al. (2003) |
| Porcine muscle (actomyosin) | DLYA | Papain | 480.5 | Arihara (2006) |
| Porcine muscle (actomyosin) | SLYA | Papain | 452.5 | Arihara (2006) |
| Porcine muscle (actomyosin) | VW | Papain | 303.4 | Arihara (2006) |
| Porcine ham skeletal muscle proteins | GKFNV, HA, LPGGGT, LPGGGHGDL | Dry-cured; Pepsin+trypsin | - | Zhu et al. (2016) |
| Porcine biceps femoris muscle proteins | GLAGA, SAGNPN | Dry-cured | - | Escudero et al. (2013) |
| Porcine fresh ground ham | QYP | Fermentation | - | Ohata et al. (2016) |
| Porcine biceps femoris muscle proteins | DLEE | Dry-cured | 504.2 | Xing et al. (2016) |
| Porcine ham muscle proteins | MDPKYR, TKYRVP | Dry-cured | - | Gallego et al. (2019) |
| Porcine longissimus dorsi muscle | VW, LW | In vitro gastro-intestinal digestion | <3,000 | Martini et al. (2019) |
| Porcine ham muscle proteins (actin) | EAGPSIVHR, ALPHAIMR, AGFAGDDAPR, VAPEEHPTL, DEAGPSIVH, AGPSIVHRK | Dry-cured | 908.1–992.1 | Wang et al. (2021) |
| Porcine ham muscle proteins (tropomyosin) | MDAIKKK, DPIIQDR | Dry-cured | 833.0–856.0 | Wang et al. (2021) |
Among the bioactive peptides, the most extensively studied are angiotensin-converting enzyme inhibitory peptides (Arihara and Ohata, 2008).
According to the World Health Organization, approximately 1.13 billion people worldwide have high blood pressure (WHO, 2013; WHO, 2021). High blood pressure can weaken the heart, damage artery walls, alter blood flow, and lead to complications such as stroke, heart disease, kidney failure, vision loss, and hardening of the arteries (Williams et al., 2018). Due to the severe side effects of various synthetic drugs used to treat hypertension, there has been extensive research on bioactive peptides derived from food proteins that can effectively treat hypertension without causing adverse reactions (Toldrá et al., 2018). Table 2 displays the antihypertensive functional peptides derived from porcine proteins, with most originating from fibrillar proteins such as myosin, actin, and troponin. Previous studies have shown that peptides with a molecular weight of less than 10 kDa have superior antioxidant and antihypertensive properties compared to larger peptides with relatively larger molecular weights. Some peptides obtained from pork proteins through in vitro digestion have shown ACE inhibitory activity (Escudero et al., 2010a; Escudero et al., 2012). For example, peptides (MYPGIA and VIPEL) derived from pork actin and GAPDH, and peptides (KRVITY and VKAGF) isolated from pork myosin heavy chain and actin exhibit ACE inhibitory activity (Escudero et al., 2010a; Muguruma et al., 2009). Peptides KAPVA and PTPVP from titin, and peptide RPR from neblin in pork enzymatic hydrolysate, show strong ACE inhibitory activity (Escudero et al., 2012). Furthermore, differences in the amino acid composition of bioactive peptides may affect ACE-inhibitory activity. For example, differences in the composition of amino acids that make up peptides, such as acidic amino acids (aspartic acid and glutamic acid), and the presence of positively charged amino acids in the carboxyl group can affect the increase in ACE-inhibitory activity (Daskaya-Dikmen et al., 2017; Peighambardoust et al., 2021).
| Protein source | Peptide sequence | Treatment | MW (Da)1) | Reference |
|---|---|---|---|---|
| Porcine muscle (myosin) | MNPPK | Thermolysin | 585.72) | Nakashima et al. (2002) |
| Porcine muscle (myosin) | ITTNP | Thermolysin | - | Nakashima et al. (2002) |
| Porcine muscle (myosin light chain) | VKKVLGNP | Pepsin | 854.0 | Katayama et al. (2007) |
| Porcine muscle (troponin) | KRQKYDI | Pepsin | 950.12) | Muguruma et al. (2009) |
| Porcine muscle (myosin heavy chain) | KRVITY | Pepsin | 805.97 | Muguruma et al. (2009) |
| Porcine muscle (actin) | VKAGF | Pepsin | 520.62 | Muguruma et al. (2009) |
| Porcine muscle (nebulin) | RPR | Pepsin+pancreatin | - | Escudero et al. (2012) |
| Porcine muscle (titin) | KAPVA | Pepsin+pancreatin | - | Escudero et al. (2012) |
| Porcine muscle (titin) | PTPVP | Pepsin+pancreatin | - | Escudero et al. (2012) |
| Porcine muscle (actin) | MYPGIA | Pepsin+pancreatin | - | Escudero et al. (2010b) |
| Porcine muscle (GAPDH) | VIPEL | Pepsin+pancreatin | - | Escudero et al. (2010b) |
| Porcine longissimus dorsi muscle (actin) | VFPS, LKYPI, AVF, MYPGIA | In vitro gastro-intestinal digestion | <3,000 | Martini et al. (2019) |
| Porcine longissimus dorsi muscle | VW, IW, VF, WL, LW, VIP, LGI, LPF, IVP, IL, LLF, WM, FIV, LR, ILP, VLP, PL, LF, IAIP, IR, IF, GLx, AV, AI, DL, NIIPA | In vitro gastro-intestinal digestion | <3,000 | Martini et al. (2019) |
| Porcine ham muscle proteins | GGVPGG, TKYRVP, HCNKKYRSEM | Dry-cured | - | Gallego et al. (2019) |
| Porcine ham muscle proteins | EL, EV, RL, EEL, ESV | Dry-cured | - | Hao et al. (2020) |
| Porcine ham muscle proteins | GA, VF | Dry-cured | - | Heres et al. (2021b), Heres et al. (2022) |
Conclusion
In this study, we presented a protein digestion analysis method and a peptide bioactivity analysis method that can be utilized for peptide acquisition. The digestive enzymes present in the intestinal tract include pepsin, trypsin, chymotrypsin, and procarboxypeptidase, with cholecystokinin and secretin playing auxiliary roles in protein digestion. Proteins are hydrolyzed in the body to generate peptides. Methods such as BCA assay, SDS-PAGE, and chromatography have been used to analyze protein digestibility and molecular weight distribution, which are applicable to peptide acquisition. In recent years, in vitro digestion simulation systems have been utilized to evaluate protein digestibility and changes in activity. In addition, the ACE inhibitory and antioxidant properties of bioactive peptides derived from pork suggest potential industrial applications. In particular, papain has been primarily used as a hydrolyzing agent for antioxidant peptides in pork. Actomyosin and tropomyosin are found in myofibrillar proteins, and they have molecular weights below 1 kDa. The antihypertensive activity is often attributed to the use of pepsin as a hydrolyzing agent in pork, with most peptides identified having a molecular weight of lower than 1 kDa. Therefore, this study can serve as a basis for the effective utilization in the development of pork-derived bioactive peptides and exploration of their bioactivity in the future. Furthermore, the advancement of pork-derived bioactive peptides may aid in promoting domestic pork consumption.